Cellular Signalling: A novel site comes into sight
- Chinese Academy of Sciences, China
- Keck School of Medicine of the University of Southern California, United States
The Hedgehog signalling pathway plays a central role in development and the maintenance of normal tissue function in adults. It has also been implicated in degenerative diseases and a multitude of cancers. A great deal of effort has been invested in designing drugs that target this pathway (Dellovade et al., 2006; Rubin and de Sauvage, 2006), and after more than a decade of work in both academic laboratories and pharmaceutical companies, several anticancer drugs that regulate the Hedgehog pathway have now entered the clinic. Most of these drugs target a membrane protein called Smoothened.
Now, in eLife, Rajat Rohatgi at Stanford University School of Medicine and co-workers in the US, UK and Singapore—including Sigrid Nachtergaele (Stanford) and Daniel Whalen (Oxford) as joint first authors—provide new insights into the regulation of Smoothened by oxysterols, a class of naturally occurring compounds, and suggest new options for the pharmacological modulation of Hedgehog signalling (Nachtergaele et al., 2013). Similar results have also been reported in Nature Chemical Biology by Daniel Nedelcu of Harvard Medical School and co-workers (Nedelcu et al., 2013), and in Developmental Cell by Benjamin Myers of Stanford University School of Medicine and co-workers (Myers et al., 2013).
Hedgehog signalling at the cell membrane involves two essential components: Patched1, which is a receptor for the Hedgehog signalling molecule, and Smoothened. In the absence of Hedgehog, Patched1 inhibits Smoothened. However, when Hedeghog binds to Patched1, Smoothened gets activated (Briscoe and Therond, 2013). There is now a general consensus that the interaction between Patched1 and Smoothened is mediated by a small molecule (or molecules) that is regulated by Patched1, rather than by direct physical interactions between the two (Taipale et al., 2002). This might explain why Smoothened has emerged as the most ‘druggable’ target in the Hedgehog pathway (Rubin and de Sauvage, 2006).
To date, the best candidates to emerge as the endogenous regulators of Smoothened activity are oxysterols, which are naturally occurring compounds produced through the oxidation of cholesterol (Corcoran and Scott, 2006; Dwyer et al., 2007). Of these, one of the most potent is 20(S)-hydroxycholesterol (20(S)-OHC). Previous work has shown that oxysterols modulate Smoothened in an allosteric manner; that is, in contrast to several other known modulators of this protein, oxysterols bind at a distinct site (Nachtergaele et al., 2012). The three new papers give detailed insights into the mechanism of action of oxysterol, and lend support to a model based on allosteric interactions.
Figure 1
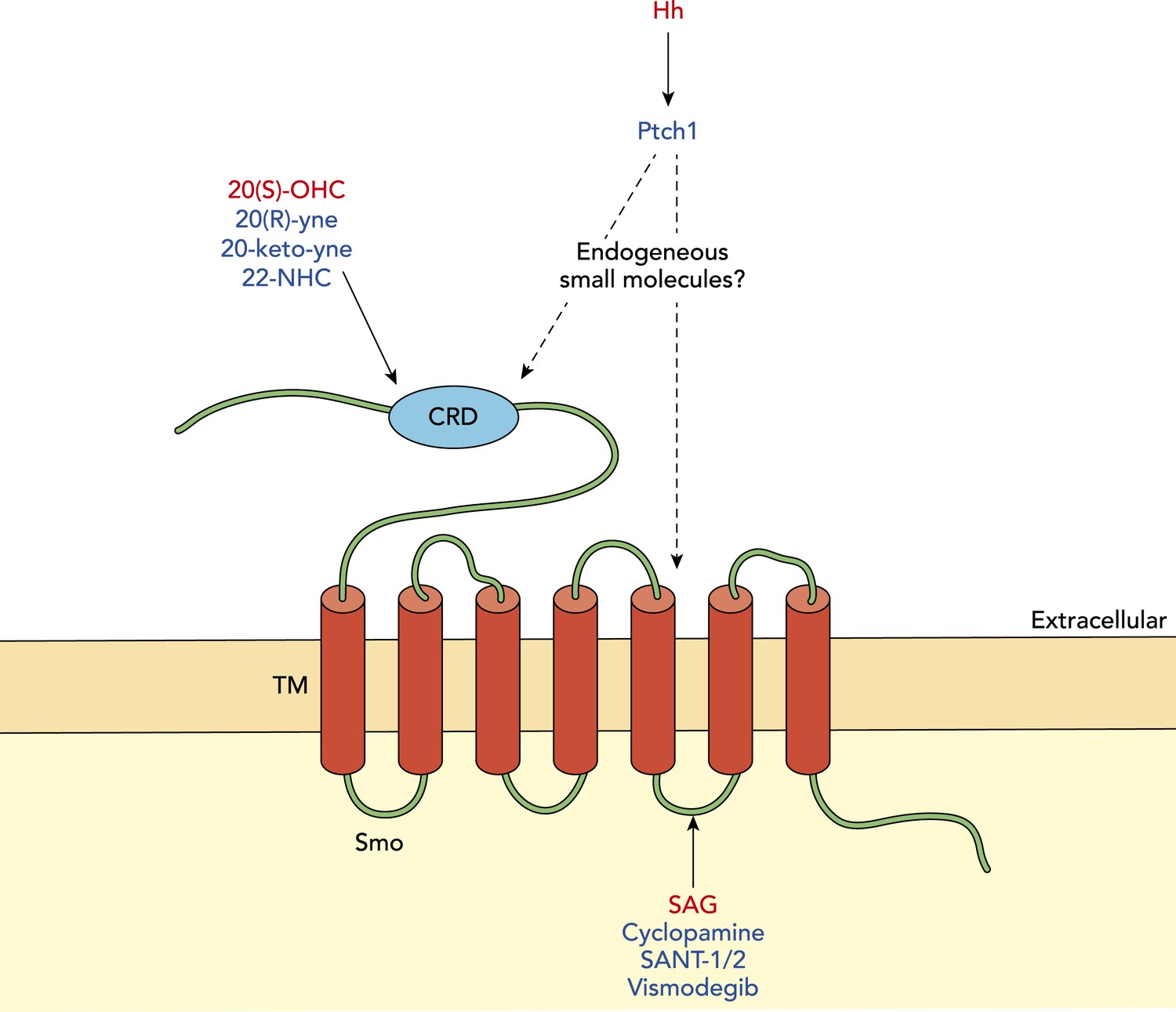
Modulation of Smoothened in vertebrates.
The signalling molecule Hedgehog (Hh) acts via a receptor called Patched1 (Ptch1) to modulate the activity of another membrane protein called Smoothened (Smo). Activators of Smo activity are shown in red and inhibitors are shown in blue. Oxysterols (such as 20(S)-OHC) can activate Smo by binding its cysteine-rich domain (CRD), while various synthetic molecules (such as 20(R)-yne, 20-keto-yne and 22-NHC) can inhibit Smo by binding the CRD; the small molecule Smoothened agonist (SAG) can increase Smo activity by interacting with its membrane-bound region, while cyclopamine and other chemicals (shown in blue) can decrease Smo activity by interacting with this region. The endogenous small molecule(s) that mediate the inhibition of Smoothened by Patched1 remain unknown.
Smoothened shares several structural features with a family of receptors called Frizzled, which bind a signalling molecule called Wnt. In particular, both possess a cysteine-rich domain at their N-terminus. The fact that this region of Frizzled binds a palmitoyl group that is present on Wnt ligands (Janda et al., 2012) suggested that oxysterols may likewise interact with the cysteine-rich domain in Smoothened; this has now been confirmed experimentally by the latest work from the groups at Stanford and Harvard. A variety of small molecules, including clinically relevant compounds that are known to interact with a cavity embedded in the transmembrane part of the protein (Wang et al., 2013), did not compete with 20(S)-OHC for binding Smoothened. Thus, it is clear that there are separate binding events for oxysterols and these other compounds.
With the aid of the crystal structure of the cysteine-rich domain in zebrafish Smoothened, Nachtergaele et al. used computational modelling to map oxysterol binding to a hydrophobic groove on the surface of this domain. This interaction had been predicted by Myers et al. and Nedelcu et al. using in silico modelling. The cysteine-rich domain is sufficient for binding oxysterol, but this property is lost when key amino acid residues within the domain are mutated.
The initiation of Hedgehog signalling depends on Smoothened building up in a cellular structure called the primary cilium (Goetz and Anderson, 2010). The two Stanford groups and the Harvard group find that Smoothened variants with a mutated oxysterol binding site show decreased translocation to the primary cilium and decreased pathway activity in response to 20(S)-OHC. Furthermore, mutant versions of Smoothened that lack the cysteine-rich domain do not respond to 20(S)-OHC, but show a response that is similar to the response of wild-type Smoothened to molecules that interact with the transmembrane domain. Together the data provide compelling evidence that oxysterols interact with the cysteine-rich domain to modulate Smoothened activity.
These findings raise two key questions: is 20(S)-OHC the endogenous small molecule that mediates the regulation of Smoothened by Patched1? And is this the major regulatory input controlling Smoothened activity?
The mutant lacking the cysteine-rich domain exhibits a higher basal level of signalling than intact Smoothened, suggesting that the cysteine-rich domain suppresses Smoothened activity (Aanstad et al., 2009; Myers et al., 2013; Nachtergaele et al., 2013; Nedelcu et al., 2013). However, when Patched1 and the mutant version of Smoothened are expressed in cells together, the basal activity of the mutant is diminished (Myers et al., 2013). This indicates that the mutant version is still subject to regulation by Patched1. Thus the cysteine-rich domain may provide tonic control of Smoothened activity, whereas Patched1-dependent processes regulate Smoothened via a site separate from the domain. Myers et al. also discovered that a cell line that responds to Hedgehog contains no endogenous 20(S)-OHC, suggesting that this oxysterol may not be essential for the regulation of Smoothened. Moreover, the same oxysterols do not bind Smoothened in Drosophila, highlighting an evolutionary divergence in regulation of the Hedgehog pathway within the animal kingdom (Nachtergaele et al., 2013; Nedelcu et al., 2013).
Depleting sterols in cells diminishes the accumulation of Smoothened in the primary cilium, and activation of the Hedgehog pathway. The observation that the Smoothened lacking the cysteine-rich domain is still sensitive to sterol depletion suggests that these compounds may have additional roles mediated through interactions outside the domain (Myers et al., 2013). The fact that the response to Hedgehog is dampened in mutants suggests that, in vertebrates, the binding of oxysterols to the cysteine-rich domain is required for maximal activity of Smoothened (Aanstad et al., 2009; Myers et al., 2013; Nachtergaele et al., 2013; Nedelcu et al., 2013).
Naturally occurring small molecule regulators of biochemical pathways can be used to guide the development of synthetic compounds that modulate the same pathways. For example, cyclopamine, a Smoothened antagonist produced by certain plants, inspired the development of the experimental anticancer drug, saridegib (Tremblay et al., 2009). The studies discussed here reveal several oxysterol analogues that inhibit the activity of the Hedgehog pathway by competing with 20(S)-OHC for access to the cysteine-rich domain in Smoothened (Nachtergaele et al., 2013; Nedelcu et al., 2013). Two of these compounds inhibit drug-resistant variants of Smoothened found in cancer patients (Nachtergaele et al., 2013). This raises the possibility of designing next generation drugs based on a sterol scaffold, and of using treatment regimes consisting of multiple drugs to target distinct sites on the Smoothened protein.
The three new studies provide a detailed picture of the role of oxysterol in the regulation of Smoothend activity, though several important questions remain. If oxysterols are not the major regulator of Smoothened function, what is? Are there multiple modulators with distinct mechanisms that still await discovery? How does Patched1 control Smoothened? This does not appear to be via oxysterols, but is presumbly through a yet-to-be-identified major control factor (or factors). Answering these questions will open up still more possibilities for pharmacological control of the Hedgehog pathway.
References
-
The mechanisms of Hedgehog signalling and its roles in development and diseaseNature Reviews Molecular Cell Biology 14:416–429.https://doi.org/10.1038/nrm3598
-
Oxysterols stimulate Sonic hedgehog signal transduction and proliferation of medulloblastoma cellsProceedings of the National Academy of Sciences of the United States of America 103:8408–8413.https://doi.org/10.1073/pnas.0602852103
-
The hedgehog pathway and neurological disordersAnnual Review of Neuroscience 29:539–563.https://doi.org/10.1146/annurev.neuro.29.051605.112858
-
Oxysterols are novel activators of the hedgehog signaling pathway in pluripotent mesenchymal cellsJournal of Biological Chemistry 282:8959–8968.https://doi.org/10.1074/jbc.M611741200
-
The primary cilium: a signalling centre during vertebrate developmentNature Reviews Genetics 11:331–344.https://doi.org/10.1038/nrg2774
-
Oxysterols are allosteric activators of the oncoprotein SmoothenedNature Chemical Biology 8:211–220.https://doi.org/10.1038/nchembio.765
-
Oxysterol binding to the extracellular domain of Smoothened in Hedgehog signalingNature Chemical Biology 9:557–564.https://doi.org/10.1038/nchembio.1290
-
Targeting the Hedgehog pathway in cancerNature Reviews Drug Discovery 5:1026–1033.https://doi.org/10.1038/nrd2086
-
Discovery of a potent and orally active hedgehog pathway antagonist (IPI-926)Journal of Medicinal Chemistry 52:4400–4418.https://doi.org/10.1021/jm900305z
Article and author information
Author details
Publication history
- Version of Record published: December 3, 2013 (version 1)
Copyright
© 2013, Wang and McMahon
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 847
- views
-
- 76
- downloads
-
- 1
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Cellular Signalling: A novel site comes into sight
eLife 2:e01680.
https://doi.org/10.7554/eLife.01680
Further reading
-
- Biochemistry and Chemical Biology
Transporter research primarily relies on the canonical substrates of well-established transporters. This approach has limitations when studying transporters for the low-abundant micromolecules, such as micronutrients, and may not reveal physiological functions of the transporters. While d-serine, a trace enantiomer of serine in the circulation, was discovered as an emerging biomarker of kidney function, its transport mechanisms in the periphery remain unknown. Here, using a multi-hierarchical approach from body fluids to molecules, combining multi-omics, cell-free synthetic biochemistry, and ex vivo transport analyses, we have identified two types of renal d-serine transport systems. We revealed that the small amino acid transporter ASCT2 serves as a d-serine transporter previously uncharacterized in the kidney and discovered d-serine as a non-canonical substrate of the sodium-coupled monocarboxylate transporters (SMCTs). These two systems are physiologically complementary, but ASCT2 dominates the role in the pathological condition. Our findings not only shed light on renal d-serine transport, but also clarify the importance of non-canonical substrate transport. This study provides a framework for investigating multiple transport systems of various trace micromolecules under physiological conditions and in multifactorial diseases.
-
- Biochemistry and Chemical Biology
- Cell Biology
Mediator of ERBB2-driven Cell Motility 1 (MEMO1) is an evolutionary conserved protein implicated in many biological processes; however, its primary molecular function remains unknown. Importantly, MEMO1 is overexpressed in many types of cancer and was shown to modulate breast cancer metastasis through altered cell motility. To better understand the function of MEMO1 in cancer cells, we analyzed genetic interactions of MEMO1 using gene essentiality data from 1028 cancer cell lines and found multiple iron-related genes exhibiting genetic relationships with MEMO1. We experimentally confirmed several interactions between MEMO1 and iron-related proteins in living cells, most notably, transferrin receptor 2 (TFR2), mitoferrin-2 (SLC25A28), and the global iron response regulator IRP1 (ACO1). These interactions indicate that cells with high MEMO1 expression levels are hypersensitive to the disruptions in iron distribution. Our data also indicate that MEMO1 is involved in ferroptosis and is linked to iron supply to mitochondria. We have found that purified MEMO1 binds iron with high affinity under redox conditions mimicking intracellular environment and solved MEMO1 structures in complex with iron and copper. Our work reveals that the iron coordination mode in MEMO1 is very similar to that of iron-containing extradiol dioxygenases, which also display a similar structural fold. We conclude that MEMO1 is an iron-binding protein that modulates iron homeostasis in cancer cells.




{kind=link}