RNA Biology: Structures to the people!
- University of the Sciences, United States
- Max Planck Institute for Molecular Biomedicine, Germany
The structures of molecules often hold the key to understanding their roles in cells. Thus, when Watson and Crick proposed the double-helix structure for DNA, they immediately speculated on how DNA may replicate. Unfortunately, working out the structures of RNA molecules is challenging, and the techniques needed to do so—X-ray crystallography and NMR—are only available at a small number of locations worldwide. This has prevented many small laboratories from embracing structural work.
Instead, some researchers have started to predict the three-dimensional structures of RNA molecules based on the sequence of nucleotides in the molecule, and there is even an ‘RNA Puzzles’ competition to compare the performance of prediction algorithms (Cruz et al., 2012; Miao et al., 2015). Moreover, the discovery of large numbers of non-coding RNAs in recent years has substantially increased the need for methods that can determine the structure of RNA molecules quickly and reliably. Now, in eLife, Rhiju Das and co-workers—including Clarence Yu Cheng as first author—describe a new approach that promises to simplify the determination of RNA structures to nanometer resolution (Cheng et al., 2015).
This approach, which is called multidimensional chemical mapping, can be likened to the power of social networking. In social networks, centralized ‘hot spots’ of communication can be identified, based on how well connected they are, and in some cases these hot spots can influence, or constrain, the activity of large numbers of other individuals in the network. Similarly, the approach developed by Cheng et al., who are based at Stanford University, identifies the three-dimensional networks of interactions between nucleotides within a given RNA molecule to guide the predictions of its structure.
Interactions between nucleotides in most RNA molecules lead to secondary structures called helices, which include loops, bulged nucleotides and multi-helical junctions. In turn, these secondary structure elements influence how the molecule folds into its final three-dimensional (or tertiary) structure. Previously, Das and others have combined protocols that predict the secondary structure of RNA with three-dimensional modelling algorithms (Kladwang et al., 2011; Cruz et al., 2012; Miao et al., 2015) to generate models of the tertiary structures of RNAs. However, the relative orientations of the secondary structure elements could not be precisely defined due to a lack of experimental data on the three-dimensional structure.
Several experimental approaches can fill this gap, but many require specialized reagents or equipment, which limits their use. Just as the growth of social media depended on developments in technological infrastructure, the current progress in multidimensional chemical mapping has been made possible by the widespread availability of instruments for deep sequencing DNA. In a wry twist of fate, sequencing has become a key tool for defining structure.
The idea to use sequencing to determine RNA structures is not new (see review by Ge and Zhang, 2015), but to date the focus has been on predicting secondary structure. Recently, however, Kevin Weeks and co-workers went a step further and used correlated data between nucleotides to predict tertiary structure, although they only reported on the interactions between two nucleotides, cytosine and adenosine (Homan et al., 2014). Cheng et al. go even further by considering potentially all of the interactions between all four of the nucleotides.
Multidimensional chemical mapping combines a technique called MOHCA (short for multiplexed •OH cleavage analysis) with deep sequencing and algorithms that predict the RNA secondary and tertiary structure (Figure 1). MOHCA uses RNA molecules that contain chemically modified nucleotides, usually one per molecule, at random positions. When the RNA molecules are treated with certain chemicals, highly reactive molecules called hydroxyl radicals are produced, and these cleave the RNA molecule near the site of the modified nucleotide. This effectively marks the positions of other nucleotides that are found near the modified nucleotide in the three-dimensional structure of the RNA molecule.
Figure 1
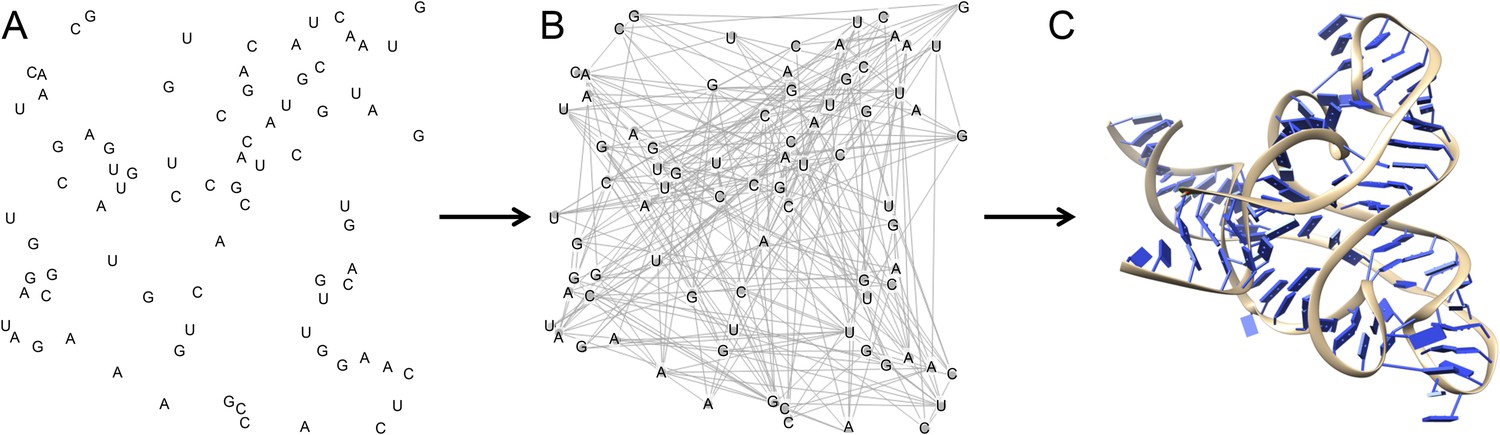
The three dimensional structure of an RNA molecule can be predicted by combining MOHCA, deep sequencing and algorithms that predict secondary and tertiary structures in the RNA.
(A) In MOHCA, copies of the RNA of interest that contain modified nucleotides—on average one per molecule—are made. These modified nucleotides produce hydroxyl groups that cleave the RNA and damage other nucleotides near to the modified nucleotide. A reverse transcriptase enzyme is used to generate DNA copies of the RNA molecules. This enzyme stops copying each RNA molecule at the point where it is cleaved or damaged. Therefore, sequencing these DNA fragments reveals the positions of nucleotides that are close to the modified nucleotide in the three-dimensional structure. This information is used to make a network of the interactions between all the nucleotides in the RNA (B). This network is then combined with algorithms that predict the secondary and tertiary structures to produce a single three-dimensional model of the tertiary structure (C). Image prepared by Nathan Baird using CodePen (codepen.io/blendmaster/full/uqibt) and structure 2YIE from the Protein Data Bank.
Previously, gel electrophoresis has been used to show where the RNA molecule is cleaved during MOHCA, but combining MOHCA with deep sequencing provides much more detailed information and significantly increases throughput. Cheng et al. constructed maps showing the interactions between the nucleotides in the molecule and combined these with algorithms that predict secondary and tertiary structures to generate a well-defined three-dimensional model of the RNA. To demonstrate the accuracy and utility of this new approach, Cheng et al. applied it to five RNAs of known structure, and to one RNA whose structure had not been released at the time. Multidimensional chemical mapping successfully predicted the three-dimensional structures of all six RNA molecules to within nanometer resolution.
However, RNA often interacts with cofactors, which remain largely undefined or otherwise make structure determination more challenging. Multidimensional chemical mapping works in solution and can help define structures in the absence of these cofactors. To test this, Cheng et al. used multidimensional chemical mapping to predict the structures of several RNAs without their cellular cofactors. Their prediction for the structure of the internal ribosomal entry site in human Hox messenger RNA will aid efforts to determine the structure of this mRNA in complex with the ribosome and other partner molecules using techniques such as cryo-electron microscopy. Cheng et al. also modeled ligand-free conformations of several riboswitches, which may guide the development of drugs that stabilize non-functional RNA conformations.
But what challenges are still ahead of us? Currently, the size of the RNA limits the analysis. Refining the modeling algorithm and the way the samples are prepared for sequencing may help to improve the predictions. A more general concern is the presence of naturally occurring modifications to RNA molecules inside cells. In highly modified structures—like transfer RNAs—these chemical groups influence the folding of the molecule. Thus, when solving the structure of an unmodified RNA molecule produced in an artificial system, an important piece in the puzzle is missing. Even when they are present, how these chemical modifications affect the cleavage of RNA by the hydroxyl radicals still needs to be assessed.
The field of structural biology is undergoing dramatic changes as improvements in technology—such as the development of free-electron lasers and improved detectors for electron microscopy—are making it possible to solve structures at atomic resolution, almost in a high-throughput manner. Will the combination of three-dimensional modeling and sequencing become a similar game changer in the field of RNA structures, being used and adapted by large numbers of researchers? Multidimensional chemical mapping clearly has the potential to transform structure determination by putting it into the hands of researchers who have experience in molecular biology and access to deep sequencing.
Today, this is almost everybody: structures to the people!
References
-
Single-molecule correlated chemical probing of RNAProceedings of the National Academy of Sciences of USA 111:13858–13863.https://doi.org/10.1073/pnas.1407306111
-
A two-dimensional mutate-and-map strategy for non-coding RNA structureNature Chemistry 3:954–962.https://doi.org/10.1038/nchem.1176
Article and author information
Author details
Publication history
- Version of Record published: July 8, 2015 (version 1)
Copyright
© 2015, Baird and Leidel
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,921
- views
-
- 169
- downloads
-
- 0
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
RNA Biology: Structures to the people!
eLife 4:e09249.
https://doi.org/10.7554/eLife.09249
Further reading
-
- Biochemistry and Chemical Biology
Transporter research primarily relies on the canonical substrates of well-established transporters. This approach has limitations when studying transporters for the low-abundant micromolecules, such as micronutrients, and may not reveal physiological functions of the transporters. While d-serine, a trace enantiomer of serine in the circulation, was discovered as an emerging biomarker of kidney function, its transport mechanisms in the periphery remain unknown. Here, using a multi-hierarchical approach from body fluids to molecules, combining multi-omics, cell-free synthetic biochemistry, and ex vivo transport analyses, we have identified two types of renal d-serine transport systems. We revealed that the small amino acid transporter ASCT2 serves as a d-serine transporter previously uncharacterized in the kidney and discovered d-serine as a non-canonical substrate of the sodium-coupled monocarboxylate transporters (SMCTs). These two systems are physiologically complementary, but ASCT2 dominates the role in the pathological condition. Our findings not only shed light on renal d-serine transport, but also clarify the importance of non-canonical substrate transport. This study provides a framework for investigating multiple transport systems of various trace micromolecules under physiological conditions and in multifactorial diseases.
-
- Biochemistry and Chemical Biology
- Cell Biology
Mediator of ERBB2-driven Cell Motility 1 (MEMO1) is an evolutionary conserved protein implicated in many biological processes; however, its primary molecular function remains unknown. Importantly, MEMO1 is overexpressed in many types of cancer and was shown to modulate breast cancer metastasis through altered cell motility. To better understand the function of MEMO1 in cancer cells, we analyzed genetic interactions of MEMO1 using gene essentiality data from 1028 cancer cell lines and found multiple iron-related genes exhibiting genetic relationships with MEMO1. We experimentally confirmed several interactions between MEMO1 and iron-related proteins in living cells, most notably, transferrin receptor 2 (TFR2), mitoferrin-2 (SLC25A28), and the global iron response regulator IRP1 (ACO1). These interactions indicate that cells with high MEMO1 expression levels are hypersensitive to the disruptions in iron distribution. Our data also indicate that MEMO1 is involved in ferroptosis and is linked to iron supply to mitochondria. We have found that purified MEMO1 binds iron with high affinity under redox conditions mimicking intracellular environment and solved MEMO1 structures in complex with iron and copper. Our work reveals that the iron coordination mode in MEMO1 is very similar to that of iron-containing extradiol dioxygenases, which also display a similar structural fold. We conclude that MEMO1 is an iron-binding protein that modulates iron homeostasis in cancer cells.




{kind=link}