Bacterial Growth: The SagA of E. faecium
An enzyme that remodels the cell wall of Enterococcus faecium helps these gut bacteria to divide and generate peptide fragments that enhance the immune response against cancer.
- Department of Genomic Medicine, Division of Cancer Medicine, The University of Texas MD Anderson Cancer Center, United States
Bacteria and other microbes residing in our intestinal tract – known as gut microbiota – are important for maintaining health and normal immunity. Among these is a group of lactic acid-producing bacteria called Enterococcus that inhibit the growth of harmful pathogens and aid digestion. As a result, these species are routinely found in probiotic supplements and fermented foods that attempt to improve gut health.
Enterococci can also become opportunistic pathogens capable of causing infections. Indeed, two of the most frequently identified species of Enterococci – E. faecium and E. faecalis – are both known to acquire antibiotic resistance. However, some strains of E. faecium have also been found to positively impact human health. For example, enrichment of E. faecium has been associated with an improved response to various types of cancer immunotherapy (Matson et al., 2018; Routy et al., 2018). Presence of these bacteria has also been shown to help prevent infections in the gut, and E. faecium are widely used as safe probiotics (Bhardwaj et al., 2010; Ishibashi et al., 2012).
E. faecium protects the gut from infections by releasing a hydrolase enzyme called secreted antigen A (SagA), which is involved in remodeling its cell wall. SagA breaks down peptidoglycans, the main component of the E. faecium cell wall, to produce small fragments called muramyldipeptides (MDPs; Kim et al., 2019). These MDPs activate receptors known as NOD2 in the host’s immune cells, leading to improved immunity in the gut.
Recent studies in mouse models showed that the MDPs generated by SagA could also increase anti-tumor immunity and improve cancer immunotherapy outcomes (Griffin et al., 2021). However, the role SagA plays in the bacteria itself remained unknown as it was believed that E. faecium needed this protein to survive. Now, in eLife, Howard Hang and colleagues – including Steven Klupt, Kyong Tkhe Fam and Xing Zhang as joint first authors – report that SagA does not affect the viability of E. faecium, but is required for cell wall remodeling and cell separation during replication (Klupt et al., 2024).
The team (who are based at Scripps Research) genetically modified E. faecium to generate a strain in which the gene for SagA was deleted. The growth of this bacterial strain was then compared to: (i) a wild-type strain, (ii) a mutant strain with inactive SagA, and (iii) a ‘complementation’ strain in which the gene for SagA had first been deleted and then re-expressed. Bacteria that lacked the gene for SagA or had an inactive version of the enzyme grew more slowly in liquid culture than wild-type E. faecium; but this was restored in the complementation strain. Experiments also showed that deleting the gene for SagA made E. faecium more sensitive to various antibiotics that target the bacterial cell wall. These exciting results raise the possibility of targeting SagA and other peptidoglycan hydrolase enzymes to overcome antibiotic resistance.
Transmission electron microscopy (TEM) – which uses a beam of electrons to generate images with ultrahigh resolution – revealed that E. faecium strains lacking the gene for SagA had more difficulty separating during replication. This caused the bacteria to cluster together, which could impair their growth. Cryo-electron tomography – a modification of TEM which can create three dimensional images of cells – was then used to quantify cell morphology parameters such as thickness of the cell wall and septum (a transient structure which helps to separate dividing cells). This revealed that deleting the gene for SagA alters the placement and projection angle of the new cell wall; however, this morphology was restored in the complementation strain.
To investigate the functional implications of deleting the gene for SagA, Klupt et al. used mass spectrometry to analyze specific components of the cell wall. They found that the strain in which the gene for SagA has been deleted generated fewer MDPs than wild-type E. faecium, which led to poor NOD2 signaling. Mouse models of cancer also did not respond to immunotherapy when they were colonized with the deficient strain. Finally, Klupt et al. demonstrated that deleting the gene for SagA reduced the population of cancer-targeting immune cells within the tumor.
Taken together, the findings show that catalytically active SagA is required for cell wall remodeling and cell separation in E. faecium, and its production of MDPs is required to mount an effective anti-tumor immune response (Figure 1). Furthermore, deleting the gene that codes for SagA impairs bacterial growth and increases sensitivity to antibiotics. Similar observations have been made in other bacteria that express peptidoglycan hydrolase enzymes, albeit via different mechanisms (Frirdich and Gaynor, 2013). This suggests that these enzymes could potentially be important antibacterial targets.
Figure 1
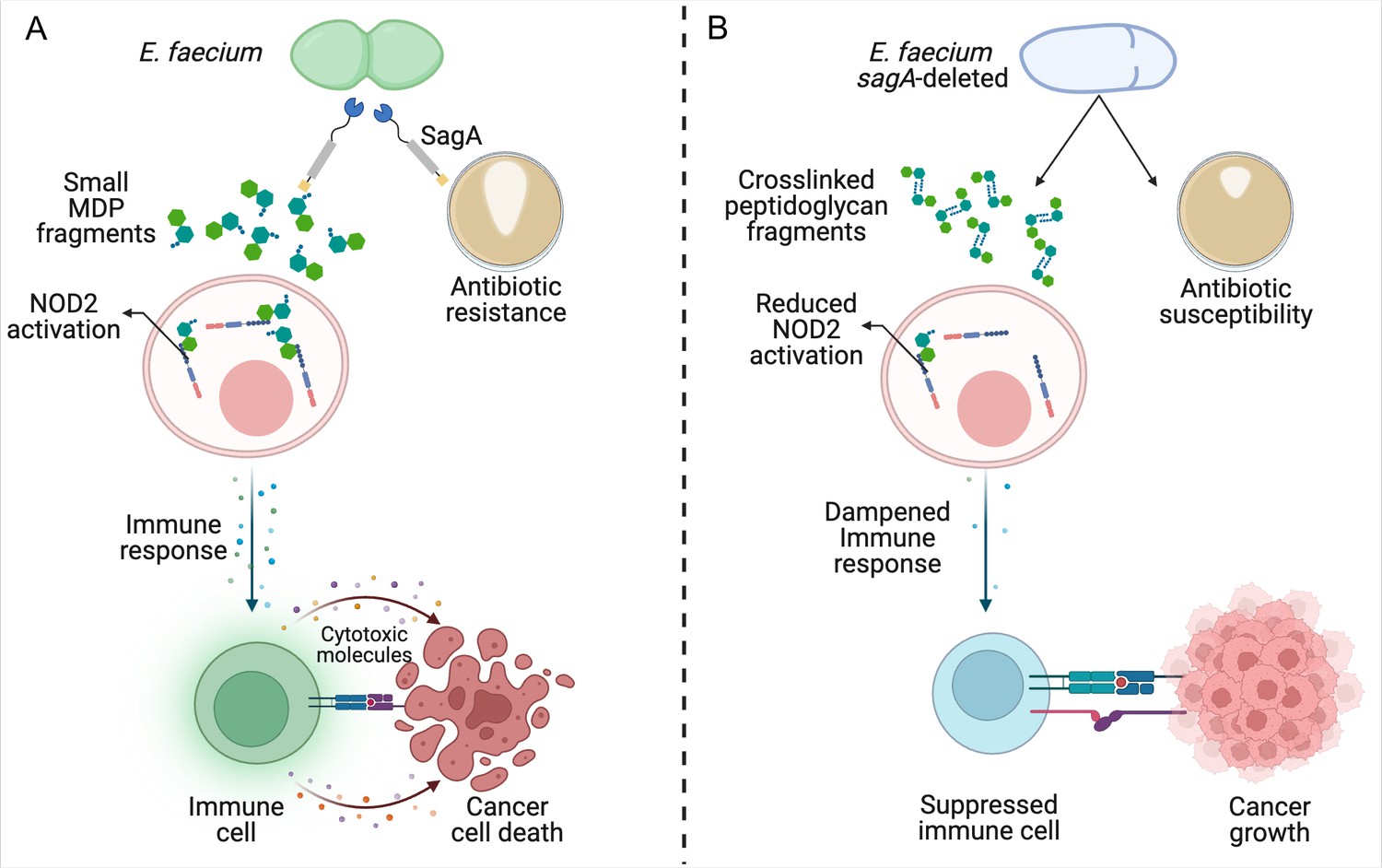
Reduced remodeling of bacterial cell walls impacts the immune response to cancer.
(A) SagA is an enzyme that helps to remodel the cell wall of E. faecium by breaking down its main component, peptidoglycan. This makes the bacteria more likely to become resistant to antibiotics (brown circle; right), allowing them to grow and form larger colonies (white shape within brown circle). SagA breaks the peptidoglycan layer into small fragments called muramyldipeptides (MDP; green hexagons), which activate NOD2 receptors in the host’s immune cells (pink). This improves the outcomes of cancer immunotherapy by triggering other cells in the immune system (green) to recognize cancer cells through receptors on the cell surface (blue and purple rectangles) and release inflammatory cytotoxic molecules that will kill them. (B) Deleting the gene for SagA (blue cell) impairs peptidoglycan remodeling and cell separation during cell division and increases the susceptibility of E. faecium to antibiotics that target the cell wall, resulting in less bacterial growth and smaller colonies. The reduced cell wall remodeling results in peptidoglycan fragments remaining crosslinked, making them too large to potently activate NOD2. This lack of sufficient NOD2 signaling prevents the immune system from mounting an appropriate immune response, leading to poorer outcomes from cancer immunotherapy.
© 2024, BioRender Inc. Figure 1 was created with BioRender.com, and published under a CC-BY-NC-ND license with permission. Further reproductions must adhere to the terms of this license.
Notably, hydrolases from other immunotherapy-promoting Enterococcus species share more than 90% sequence homology in their catalytic domain. Future studies investigating how bacteria regulate the activity of these hydrolases could lead to better treatments for cancer and combating antibiotic resistance.
References
-
Safety assessment and evaluation of probiotic potential of bacteriocinogenic Enterococcus faecium KH 24 strain under in vitro and in vivo conditionsInternational Journal of Food Microbiology 141:156–164.https://doi.org/10.1016/j.ijfoodmicro.2010.05.001
-
Peptidoglycan hydrolases, bacterial shape, and pathogenesisCurrent Opinion in Microbiology 16:767–778.https://doi.org/10.1016/j.mib.2013.09.005
-
Purification and characterization of multiple bacteriocins and an inducing peptide produced by Enterococcus faecium NKR-5-3 from Thai fermented fishBioscience, Biotechnology, and Biochemistry 76:947–953.https://doi.org/10.1271/bbb.110972
Article and author information
Author details
Publication history
- Version of Record published: April 5, 2024 (version 1)
Copyright
© 2024, Prasad and Jenq
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 424
- views
-
- 51
- downloads
-
- 0
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Bacterial Growth: The SagA of E. faecium
eLife 13:e97277.
https://doi.org/10.7554/eLife.97277
Further reading
-
- Chromosomes and Gene Expression
- Immunology and Inflammation
Ikaros is a transcriptional factor required for conventional T cell development, differentiation, and anergy. While the related factors Helios and Eos have defined roles in regulatory T cells (Treg), a role for Ikaros has not been established. To determine the function of Ikaros in the Treg lineage, we generated mice with Treg-specific deletion of the Ikaros gene (Ikzf1). We find that Ikaros cooperates with Foxp3 to establish a major portion of the Treg epigenome and transcriptome. Ikaros-deficient Treg exhibit Th1-like gene expression with abnormal production of IL-2, IFNg, TNFa, and factors involved in Wnt and Notch signaling. While Ikzf1-Treg-cko mice do not develop spontaneous autoimmunity, Ikaros-deficient Treg are unable to control conventional T cell-mediated immune pathology in response to TCR and inflammatory stimuli in models of IBD and organ transplantation. These studies establish Ikaros as a core factor required in Treg for tolerance and the control of inflammatory immune responses.
-
- Evolutionary Biology
- Immunology and Inflammation
CD4+ T cell activation is driven by five-module receptor complexes. The T cell receptor (TCR) is the receptor module that binds composite surfaces of peptide antigens embedded within MHCII molecules (pMHCII). It associates with three signaling modules (CD3γε, CD3δε, and CD3ζζ) to form TCR-CD3 complexes. CD4 is the coreceptor module. It reciprocally associates with TCR-CD3-pMHCII assemblies on the outside of a CD4+ T cells and with the Src kinase, LCK, on the inside. Previously, we reported that the CD4 transmembrane GGXXG and cytoplasmic juxtamembrane (C/F)CV+C motifs found in eutherian (placental mammal) CD4 have constituent residues that evolved under purifying selection (Lee et al., 2022). Expressing mutants of these motifs together in T cell hybridomas increased CD4-LCK association but reduced CD3ζ, ZAP70, and PLCγ1 phosphorylation levels, as well as IL-2 production, in response to agonist pMHCII. Because these mutants preferentially localized CD4-LCK pairs to non-raft membrane fractions, one explanation for our results was that they impaired proximal signaling by sequestering LCK away from TCR-CD3. An alternative hypothesis is that the mutations directly impacted signaling because the motifs normally play an LCK-independent role in signaling. The goal of this study was to discriminate between these possibilities. Using T cell hybridomas, our results indicate that: intracellular CD4-LCK interactions are not necessary for pMHCII-specific signal initiation; the GGXXG and (C/F)CV+C motifs are key determinants of CD4-mediated pMHCII-specific signal amplification; the GGXXG and (C/F)CV+C motifs exert their functions independently of direct CD4-LCK association. These data provide a mechanistic explanation for why residues within these motifs are under purifying selection in jawed vertebrates. The results are also important to consider for biomimetic engineering of synthetic receptors.




{kind=link}